

今回の心電図に関わるテーマは「活動電位」です。
うーん・・・参考書の活動電位やらイオン流入出のページを見ただけで本を閉じたくなるようなところです。というわけで、今回は非常に眠たくなるような内容です。
執筆するにあたって、持っている参考書を何冊も比較し、バラバラに記載されていることを何とかまとめようと頑張りました。
理解することと、覚え切ることに時間がかかりそうな内容ですが、ここを理解してしまえば、心電図がより楽しくなる気がします。頑張りましょう。
膜電位とは
まずは、活動電位からおさらいです。
活動電位とは、神経細胞の細胞膜を挟んで、細胞内と細胞外で生じる電位差のことです。
この電位差が生じる原因というのは、細胞内外のイオン分布の差、つまりイオン濃度の差によって発生します。
膜電位に関わるイオン
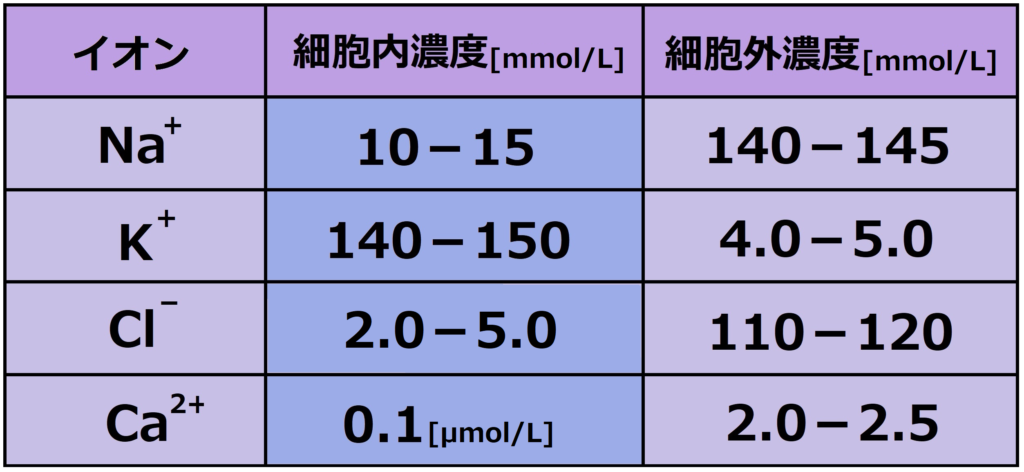
後述する静止膜電位や活動電位に関連するイオンは、Na+、K+、Ca2+、Cl–の4種類です。
ここで、細胞内外の上記イオンの濃度を確認しておきます。
細胞内のイオン濃度は、普段の採血で見るデータと全然異なることに注目ですね。
特にNa+、K+、ついでにCa2+ですね。
細胞膜の構造
次に細胞膜の構造についておさらいです。
いやー、懐かしいですね。高校時代は生物選択でしたので覚えたものです。
細胞膜は脂質二重膜構造になっています。何が二重構造なのかといいますと、2分子のリン脂質が親水部が疎水部を挟み込むように重なっている構造からこう呼ばれています。
リン脂質は頭部は親水性のリン酸、尾部は疎水性の脂肪酸の2つの成分で構成されています。
「リハスタ」リハ学生・新人セラピスト応援ブログ:生理学基礎講座|静止電位と活動電位細胞膜構造
https://rehab-rooms.com/resting_potential-action_potential/
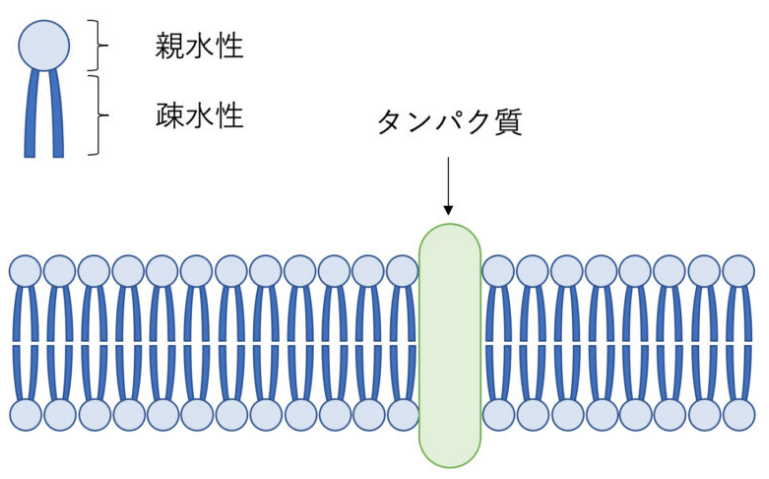
そして、この細胞膜にはトランスポータ(ポンプ、キャリア)、チャネルといった膜タンパク質が埋め込まれています。この膜タンパク質とリン脂質は流動的に動くことが可能で、これを流動モザイクモデルと言います(懐かしい!!)。
そうです、このポンプやトランスポータ、チャネルといった膜タンパク質がNa+、K+、Ca2+、Cl–などのイオンの細胞膜内外でのやり取りをしているわけです。
膜タンパク質
静止膜電位や活動電位のお話に入るまでの前置きが長くなりますがお付き合いください。
膜タンパク質の役割について、今回は特に静止膜電位や活動電位に関わる内容のみ取り上げます。
脂質二重膜構造により、細胞質とその外界は隔てられていますが、当然細胞膜は全ての物質を通すわけではなく、特定の物質だけを通します。
リン脂質二重膜は分子量が小さくて疎水性である程通過しやすいです。拡散による物質の移動はありますが、イオン、タンパク質、グルコースなどは通ることができません。
ですので、細胞膜は必要な物質を膜タンパク質を利用して通過させます。
物質によって不透過・通過の透過性が異なることを選択的透過性と言いましたね。
選択的透過性には、受動輸送と能動輸送があります。
ポンプやチャネルの話も織り交ぜて説明します。
受動輸送
受動輸送は、細胞内外での物質の濃度勾配に従うという自然の摂理に基づいた物質の移動であり、エネルギーを必要としません。つまりは拡散ですね。
この拡散にも何種類かありまして、以下に説明します。
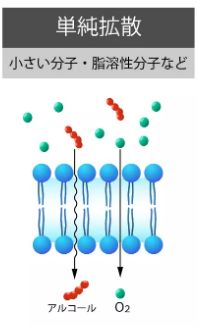
単純拡散
酸素、二酸化炭素、アルコールなど小分子の脂溶性分子は拡散により細胞膜を通過します。
イオン、タンパク質、グルコース、アミノ酸などはリン脂質部分を透過できません。
受験のミカタ「細胞膜の構造と働きを徹底解説!受動輸送・能動輸送をマスターしよう」単純拡散
https://juken-mikata.net/how-to/biology/cell-membrane.html
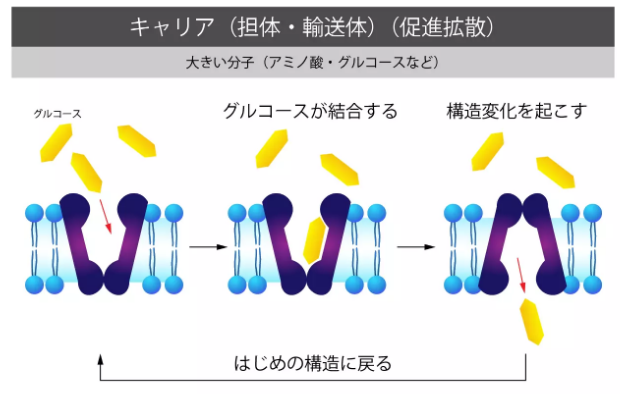
キャリア(促進拡散)
タンパク質、グルコース、アミノ酸など大分子物質は、キャリア(担体、輸送体)と呼ばれる膜タンパク質によって細胞膜を通過します。
受験のミカタ「細胞膜の構造と働きを徹底解説!受動輸送・能動輸送をマスターしよう」キャリア
https://juken-mikata.net/how-to/biology/cell-membrane.html
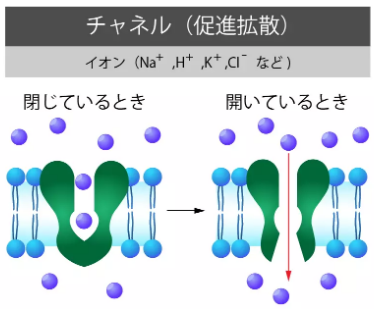
チャネル(促進拡散)
水分子はアクアポリン、イオンはイオンチャネルによって細胞膜を通過します。
受験のミカタ「細胞膜の構造と働きを徹底解説!受動輸送・能動輸送をマスターしよう」チャネル
https://juken-mikata.net/how-to/biology/cell-membrane.html
能動輸送
能動輸送は受動輸送と異なり、エネルギーを使用して物理法則に反した物質の移動を示します。
能動輸送に必要なエネルギーは基本的にはATPです。
能動輸送で使用される膜タンパク質をポンプと呼ばれ、有名などころと言えばNa+/K+ ATPase(Na+/K+ ATPアーゼ)というもので、いわゆる「ナトリウム-カリウムポンプ」のことです。
受験のミカタ「細胞膜の構造と働きを徹底解説!受動輸送・能動輸送をマスターしよう」受動輸送
https://juken-mikata.net/how-to/biology/cell-membrane.html

静止膜電位
前置きが長くなりましたが、ここから本題に入ります。
細胞が興奮していない定常状態では、膜電位は陰性に帯電しており、心筋で-90[mV]、洞結節や房室結節では-60[mV]です。
では、どうして陰性に帯電しているかというと、①Na+/K+ ATPaseの働き、②K+の濃度勾配維持(K+平衡電位)の働きにあります。それぞれ説明します。
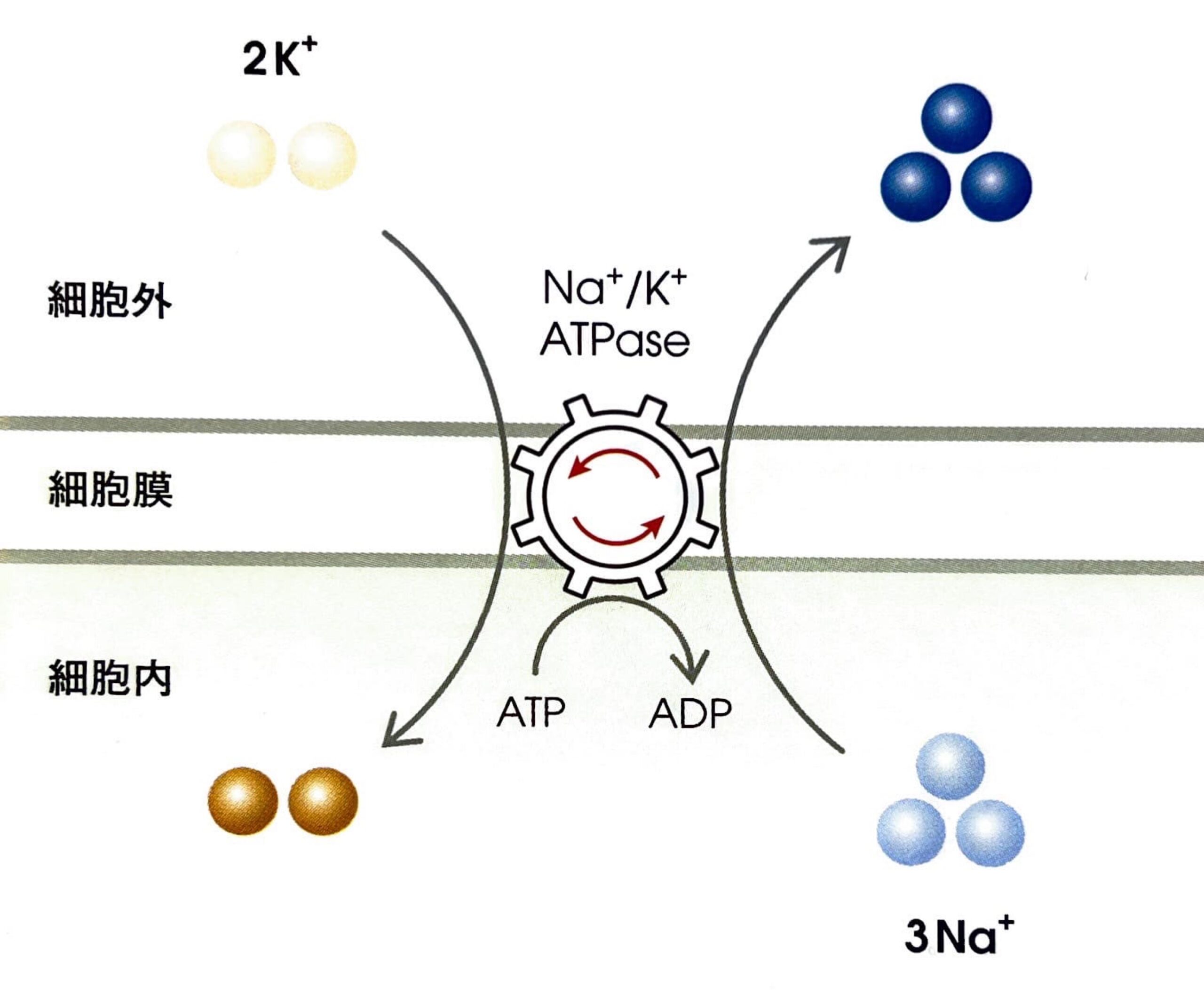
①Na+/K+ ATPase
細胞内はK+濃度が高く、細胞外はNa+濃度が高くなっているのですが、まさにこれがNa+/K+ ATPaseの働きによって引き起こされている現象です。
Na+/K+ ATPaseは1分子のATPを加水分解して、分解によって得られたエネルギーで3分子のNa+を細胞外へ、2分子のK+を細胞内へ能動輸送しています。
つまり、電荷のバランス的には細胞外へ+電荷が多く出されています。逆のことを言えば、細胞内は陰性に帯電しているということです。
これが、細胞内は陰性、細胞外は陽性に帯電している証明になります。
しかし、このNa+/K+ ATPaseの働きが静止膜電位が陰性である直接的な原因ではありません。
次のK+濃度勾配の話で明らかになります。
大谷 修, 堀尾 嘉幸「カラー図解 人体の正常構造と機能II 循環器」, p28Na+/K+ ATPaseの働き
②K+平衡電位
細胞内外でNa+、K+、Cl–の各イオンは濃度差が異なることは周知の事実ですが、心筋細胞が興奮していない定常状態では、Na+とCl–の膜のイオン透過性は非常に低くなっています。
一方で、K+に関しては内向き整流性K+チャネルや漏洩K+チャネルというチャネルが存在しているためにK+のイオン透過性は高くなっています。
漏洩K+チャネルによりK+は常に細胞の外に流れ出て平衡を保とうとしているのですが、逆にK+流出を引き留めようとする負の電位が発生しており、この電位がK+の濃度勾配と釣り合っているのです。
この電位をK+平衡電位(EK)と呼ばれます。
K+平衡電位の発生が静止膜電位の-90[mV]を示しています。
静止膜電位はK+の濃度勾配で生じているのです。
Na+にも同じことが生じそうですが、細胞膜にはNa+を通す膜タンパク質がほとんどないのでNa+の濃度勾配は膜電位に関与しません。
参考書や高校生物の教科書を見ても、静止膜電位が-70[mV]とされているのも良く見かけます。
もしかすると、心筋ではなく「ニューロン」として大きなくくりで表されているからかもしれません。
完全に参考程度の内容ですが、K+の平衡電位 EKはNernstの式で算出できます。
※ただし、R:気体定数、T:絶対温度、F:ファラデー定数、Zk:K+の原子価(+1)、
[K+]o:細胞外K+濃度、[K+]i:細胞内K+濃度、37℃でRT/FZk=61.5[mV]、
[K+]o=5[mMol/L]、[K+]i=140[mMol/L]とする。
\begin{align}
E_K[mV]&=\frac{RT}{FZ_K}{ln}\frac{[K^+]_o}{[K^+]_i}&\\
&=2.3\frac{RT}{FZ_K}{log_{10}}\frac{[K^+]_o}{[K^+]_i}&\\
&≒-89&
\end{align}
活動電位とは
お待たせしました。本日のメインテーマです。
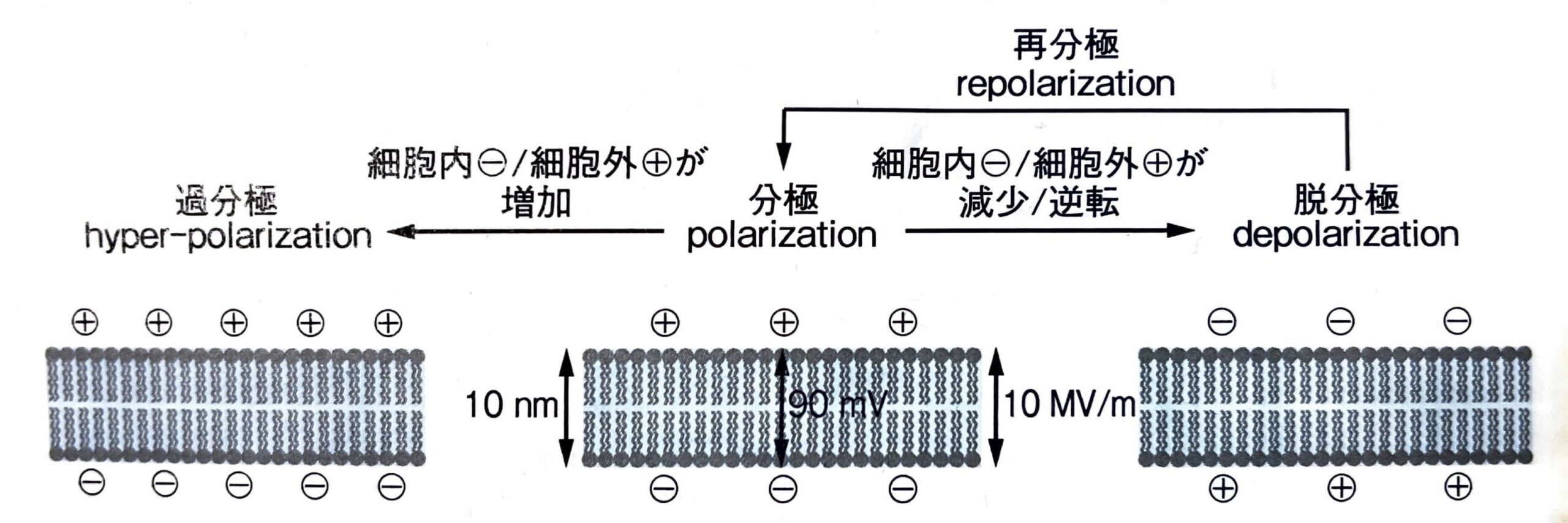
細胞膜は静止膜電位の状態から脱分極をして、再分極するサイクルを延々と続けています。
細胞膜が脱分極と再分極をするという、静止膜電位から一過性に電位が陽性になった後に元に戻る一連の膜電位の変化を活動電位と呼びます。
以下に脱分極、再分極、そして分極そのものの説明をします。
- 分極
-
静止膜電位の状態で、細胞内が陰性、細胞外が陽性に分かれること。
- 脱分極
-
静止膜電位状態の細胞が刺激を受け、一過性に電位がゼロに近づく現象のこと。
その名の通り、分極状態から脱するという意味。
オーバーシュートで一瞬陽性に傾く。 - 再分極
-
脱分極した状態から、再度静止膜電位へ戻って再度分極する現象のこと。
日本不整脈心電学会「実力心電図【改訂版】」, p17細胞膜の電気活動サイクル
活動電位の波形
活動電位は細胞膜に存在するイオンチャネルの開口と閉鎖による種々のイオンの流入出によって引き起こされています。
この活動電位は、心房筋・心室筋・His-Purkinje系の活動電位の形状と洞結節・房室結節での形状が異なるようです。
下図は心室筋の活動電位の波形、そして心室筋と洞結節及び房室結節の活動電位の波形比較です。
大谷 修, 堀尾 嘉幸「カラー図解 人体の正常構造と機能II 循環器」, p29心室筋の活動電位
大谷 修, 堀尾 嘉幸「カラー図解 人体の正常構造と機能II 循環器」, p28心室筋と洞結節及び房室結節の活動電位の波形比較
活動電位の成り立ち
心室筋の活動電位の成り立ちを説明します。
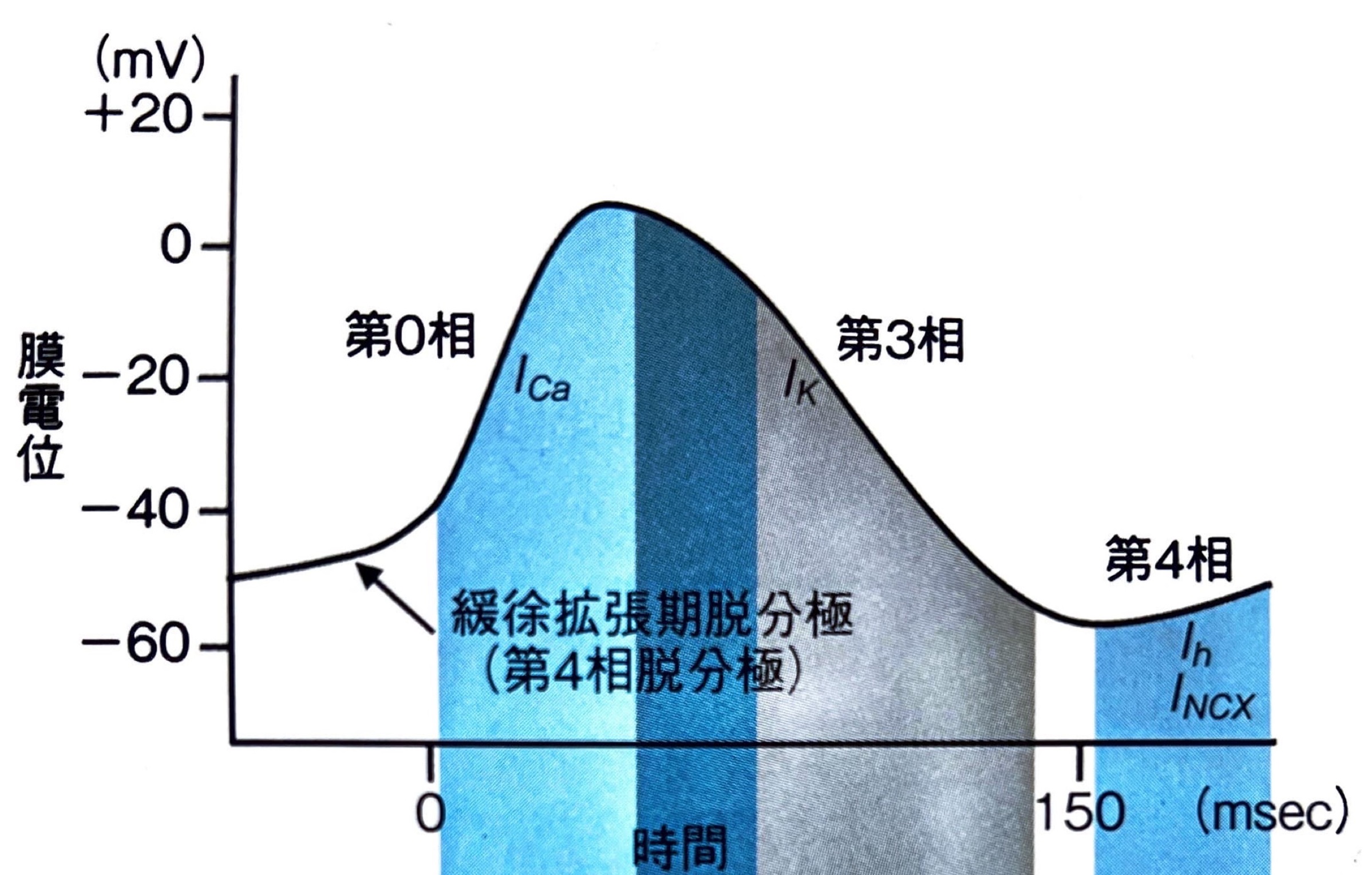
心室筋の活動電位の波形は、0相から4相までの5相から構成されます。
まずは5つの相が波形のどこに当たるのかをご確認ください。
大谷 修, 堀尾 嘉幸「カラー図解 人体の正常構造と機能II 循環器」, p30心筋細胞の活動電位波形と5つの相
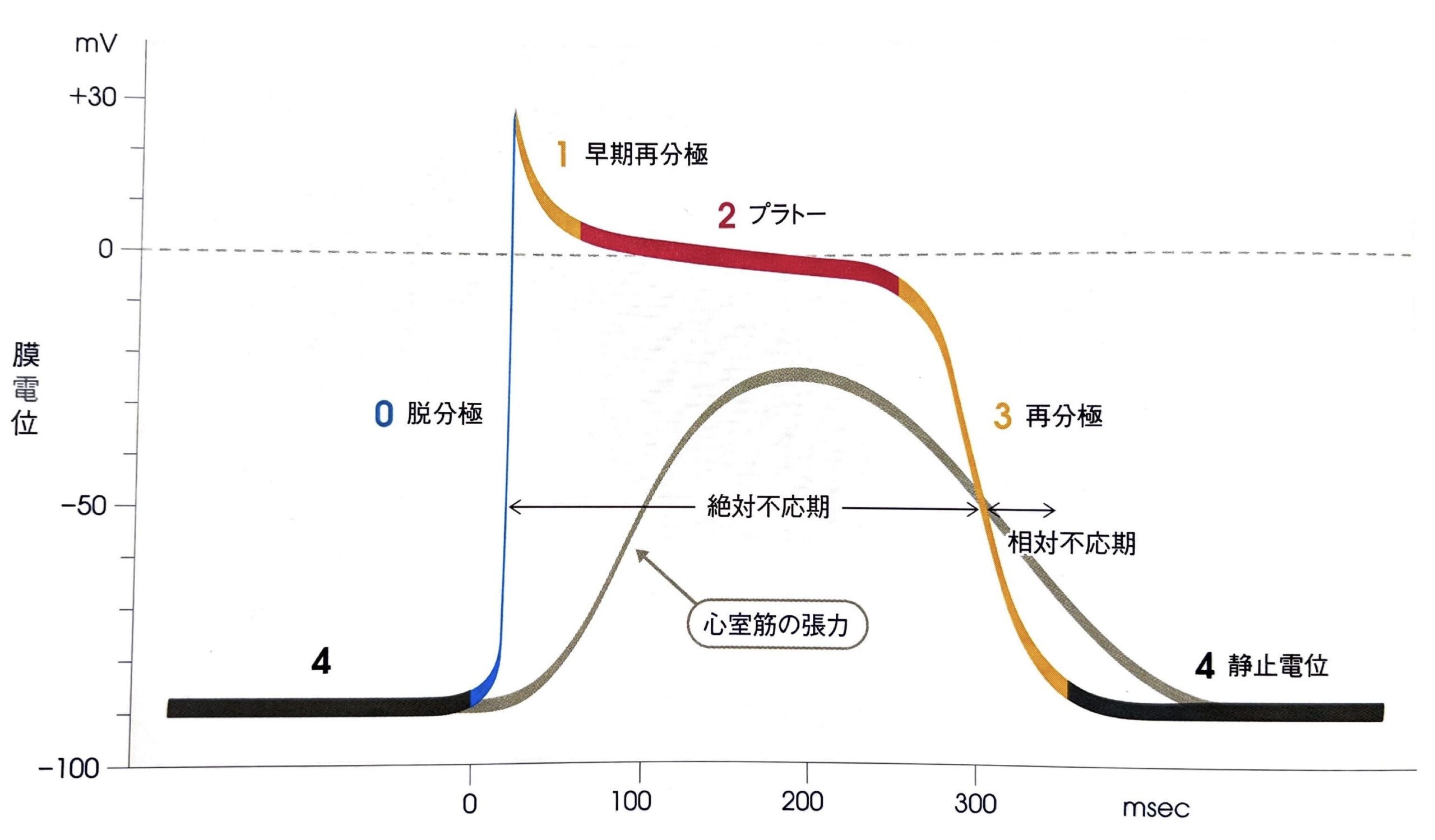
上図では、第0相と第2相が脱分極、第1相と第4相が再分極を表していることが確認できます。
では、眠い眠いそれぞれの相の説明をします。
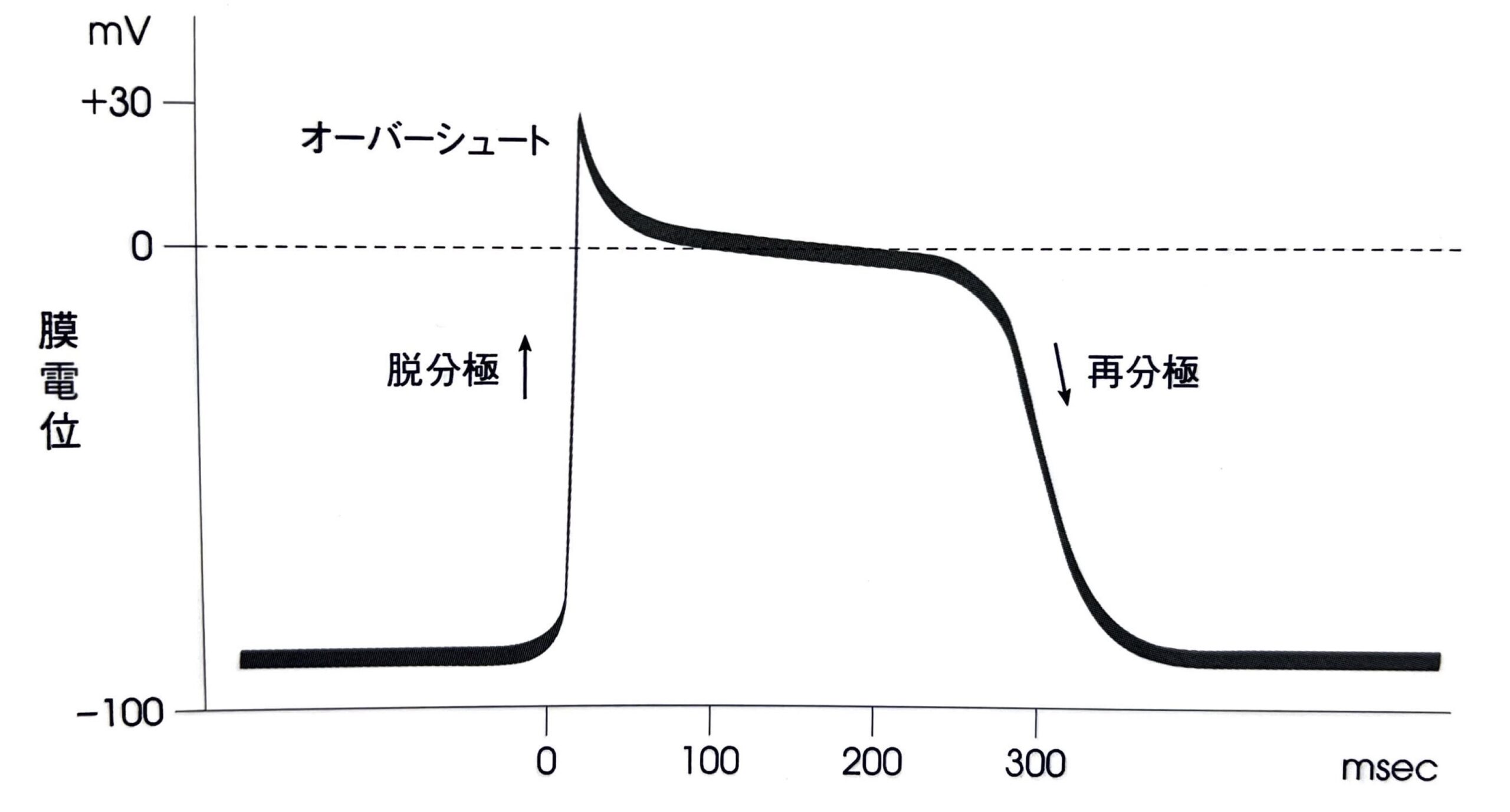
第0相(活動電位立ち上がり相)
活動電位立ち上がり相であり、電位依存性Na+チャネルの開口に伴いNa+の細胞内への流入を引き起こします。
細胞内外でNa+濃度勾配が10倍以上あり、静止膜電位が陰性であることもあり、電位依存性Na+チャネルの開口により一気にNa+が流入するために急激な立ち上がりとなります。
急激な立ち上がりのため、膜電位は+30[mV]程度までオーバーシュートします。
電位依存性Na+チャネルはすぐに閉鎖します。
大谷 修, 堀尾 嘉幸「カラー図解 人体の正常構造と機能II 循環器」, p31第0相

大谷 修, 堀尾 嘉幸「カラー図解 人体の正常構造と機能II 循環器」, p29オーバーシュート
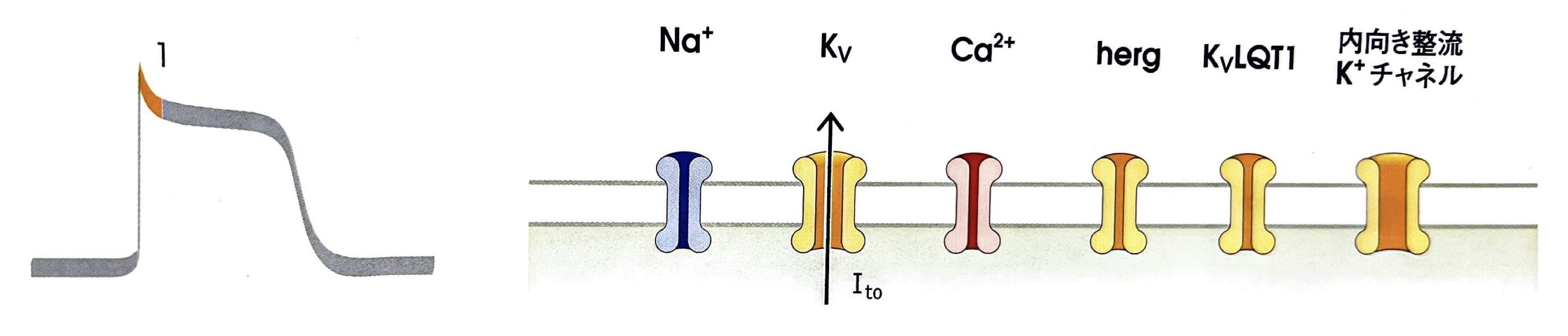
第1相(早期一過性再分極相)
電位依存性K+チャネル(KVチャネル)が開口して第1相へ移行します。
細胞外へK+が流出することで膜電位が僅かに下がります。
そして間もなく電位依存性K+チャネルが閉鎖します。
大谷 修, 堀尾 嘉幸「カラー図解 人体の正常構造と機能II 循環器」, p31第1相
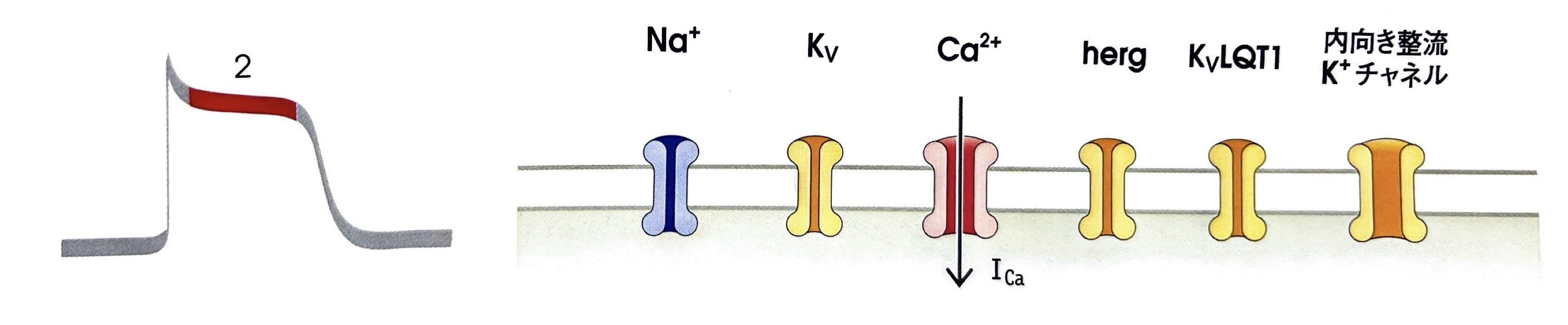
第2相(プラトー相)
L型電位依存性Ca2+チャネルの開口によりプラトー相である第2相へ移行します。
細胞内外のCa2+濃度勾配は1000倍と非常に高いため、細胞内へCa2+が流入してきます。
電位依存性Na+チャネルよりも発現時間が長いので、骨格筋の活動電位時間よりも長いのが特徴です
大谷 修, 堀尾 嘉幸「カラー図解 人体の正常構造と機能II 循環器」, p31第2相
第3相(再分極相)
再分極の過程である第3相はL型電位依存性Ca2+チャネル閉鎖と電位依存性K+チャネル群の開口により移行します。
電位依存性K+チャネル群というのは、2種類のK+チャネルが存在しており、最初に開口するhergチャネルと遅れて開口するKVLQT1があります。
この2つのK+チャネルにより細胞外へK+を流入させ、細胞内電位をK+平衡電位(EK)まで低下させます。さらに膜電位が下がってもすぐには閉鎖せずにゆっくりと閉鎖させる遅延整流性という特性を持っており、この特性により膜電位を深く落とし込むことができます。
大谷 修, 堀尾 嘉幸「カラー図解 人体の正常構造と機能II 循環器」, p31第3相

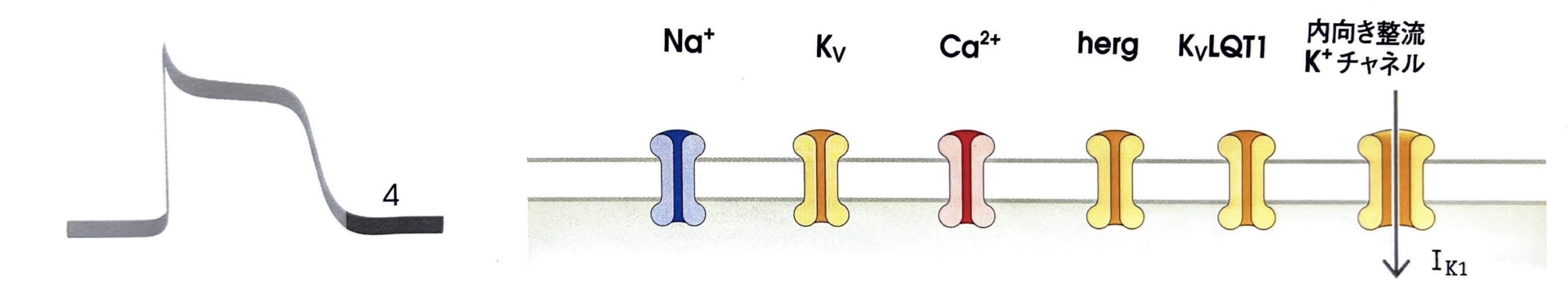
第4相(静止膜電位)
第4相は第3相で深く落とし込まれた膜電位を維持する相となります。
定常状態で常に開口している内向き整流K+チャネルが存在しており、細胞内へK+を通しやすく、細胞外へ通しにくいという性質も持ちます。そのため、常に開口していても活動電位が発生している間は、細胞外方向へK+が流出しにくいため、活動電位を阻害しません。
そしてK+平衡電位(EK)、つまり静止膜電位まで膜電位が下がってくるとK+を細胞外へ、過分極するとK+を細胞内へ通すことで静止膜電位付近に膜電位を安定させるわけです。
以上が活動電位の成り立ちとなります。
・・・まとめたことで筆者自身が理解が深まりました。
大谷 修, 堀尾 嘉幸「カラー図解 人体の正常構造と機能II 循環器」, p31第4相
活動電位に関わるイオンチャネル
各相の話をする前に、活動電位に関わるイオンチャネルについて説明を補足しておきます。
- 電位依存性Na+チャネル
第0相で開口し、Na+の細胞内への流入を引き起こす。
→電位依存性Na+電流:INa(内向き) - 電位依存性K+チャネル(KVチャネル)
第1相で開口し、K+の細胞外への流出を引き起こす。
→一過性外向きK+電流:ITO - L型電位依存性Ca2+チャネル
第2相で開口し、Ca2+の細胞内への流入を引き起こす。
→電位依存性L型Ca2+電流:ICaL(内向き) - hergチャネル(電位依存性K+チャネル herg/KV11.1)
第3相で開口し、K+の細胞外への流出を引き起こす。
→遅延整流K+電流(急速活性化K+電流):IKr(外向き)
※hERG(human ether-a-go-go related gene):ヒト急速活性型遅延整流カリウムチャネル遺伝子 - KVLQT1(電位依存性K+チャネル KVLQT1/KV7.1)
第3相で開口し、K+の細胞外への流出を引き起こす。
→遅延整流K+電流(緩徐活性化K+電流):IKS(外向き) - 内向き整流K+チャネル
第4相で開口し、K+の細胞内への流入を引き起こす。
→遅延整流K+電流(緩徐活性化K+電流):IKS(外向き)
活動電位中のイオン電流
下図は各相におけるイオン電流の図です。
2種類のグラフを参考書よりお借りして掲載させていただきます。
大谷 修, 堀尾 嘉幸「カラー図解 人体の正常構造と機能II 循環器」, p29イオン電流①

日本不整脈心電学会「実力心電図【改訂版】」, p18イオン電流②

洞結節・房室結節の活動電位
洞結節・房室結節の活動電位は心房筋・心室筋・His-Purkinje系の活動電位と性質が異なると先述しました。洞結節・房室結節についても少し触れたいと思います。
まず、心室筋との活動電位の波形比較です。
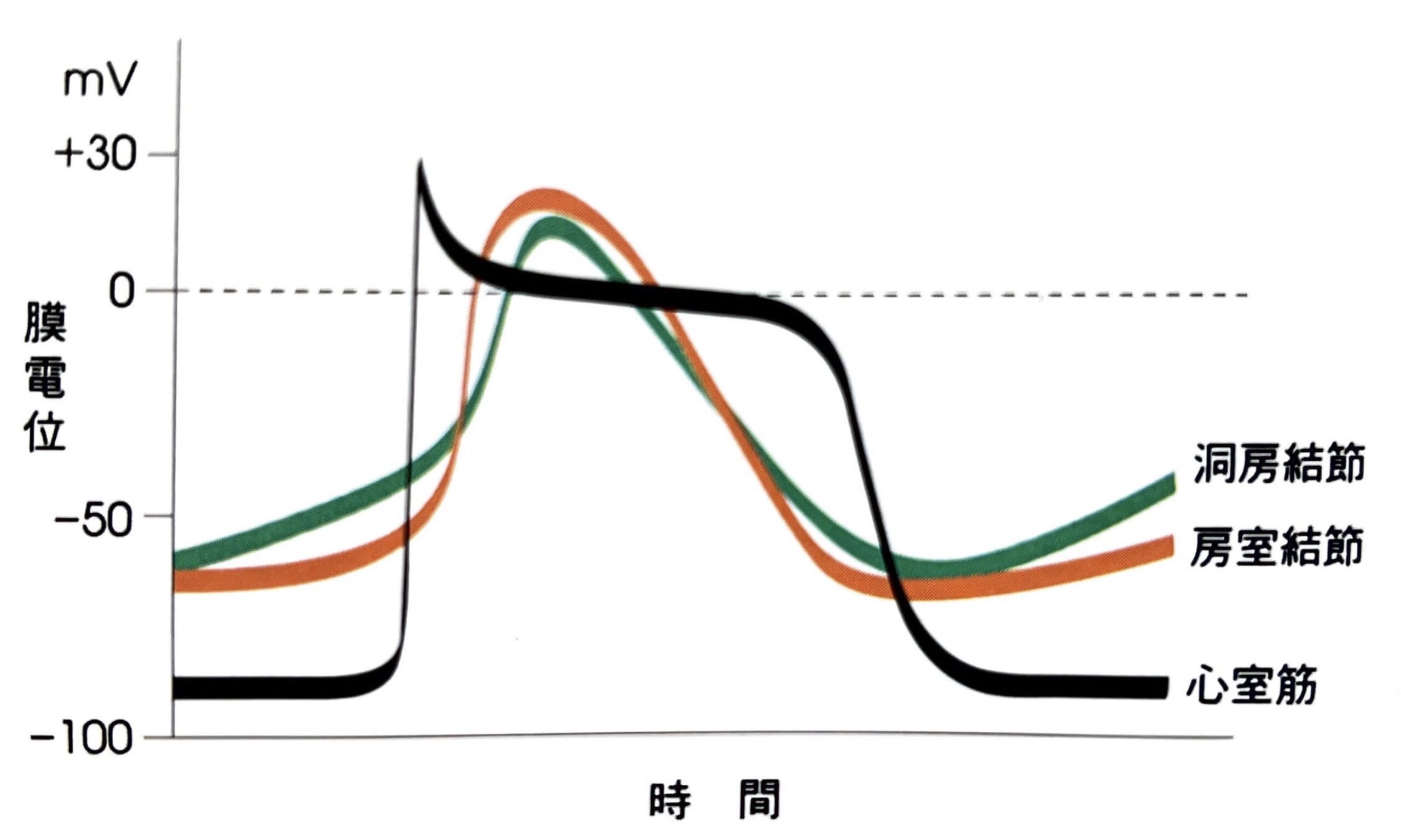
洞結節・房室結節は心室筋と比較すると、第1相と第2相が無く、第0相からいきなり第3相に移行するというという違いがあります。
日本不整脈心電学会「実力心電図【改訂版】」, p19洞結節・房室結節の活動電位波形
具体的な違いを以下に述べます。
第0相の立ち上がりが緩やか
心室細胞と比較すると第0相の立ち上がりが緩やかです。
これは洞結節・房室結節では内向き整流K+チャネルの発現が少なく、静止膜電位が-60[mV]程度となっており、電位依存性Na+チャネルが開口できなくなっています。そのため、Na+の流入による活動電位の急激な立ち上がりが発生しません。
洞結節・房室結節の活動電位は、L型電位依存性Ca2+チャネルによるCa2+の流入によるものであり、Na+電流よりも約1/100の電流密度のために立ち上がりが緩徐となっています。
Caブロッカーの投与で心拍数が遅くなり房室伝導が抑制されるのはL型電位依存性Ca2+チャネルによる活動電位であるためです。
簡単に言いますと、洞結節・房室結節はCa2+活動電位、心室はNa+活動電位です。
第4相の緩徐な脱分極
第4相は緩徐な脱分極となっており、緩徐拡張期脱分極または第4相脱分極と呼ばれています。
- 過分極活性化陽イオンチャネル(funnyチャネル、ペースメーカチャネルとも)
内向きの過分極活性化陽イオン電流(【Ih(If)】funny電流、ペースメーカ電流とも)
→細胞膜クロックと呼ばれる - Na+/Ca2+交換輸送体(NCX:Na+/Ca2 exchangers)
3つのNa+を細胞内へ、1つのCa2を細胞外への輸送に伴う内向き電流【INCX】
→カルシウムクロックと呼ばれる
浅い最大拡張期電位
洞結節・房室結節の静止膜電位及び最大拡張期電位が-60[mV]程度と浅くなっているのは、内向き整流K+チャネルの発現が少ないために、K+の透過性が低く、K+の移動が制限されているためです。
そのため第3相の電位依存性K+チャネル群による遅延整流K+電流が再分極と静止膜電位の維持を担っているわけです。
日本不整脈心電学会「実力心電図【改訂版】」, p19洞結節・房室結節の活動電位とイオン電流

不応期
細胞は閾値以上の刺激があった場合、刺激の強さに関係なく一定の活動電位の反応を示す特性を持っています。これを全か無かの法則と言いました。
しかし、活動電位発生中では細胞は興奮性を失っており、刺激を与えても活動電位が発生しないという期間があります。この時期を不応期と言います。
絶対不応期
特に第0相、第1相、第2相、そして第3相の前半までは、活動電位発生中は刺激が与えられても興奮はできず、この期間を絶対不応期(absolute refractory period)と言います。
電位依存性Na+チャネルは-65[mV]で開口します。そのため、電位差が-60[mV]程度まで落ち込まないと心筋細胞はいかなる刺激にも反応を示すことはありません。
相対不応期
第3相後半では電位差が-60[mV]以下となってくるため、電位依存性Na+チャネルの不活化から回復を見せ始めます。刺激に対して反応可能なチャネルが徐々に増えてきてますので、この時期を相対不応期と言います。
心電図上では、絶対不応期から相対不応期の移行部がT波頂上付近となります。
この時期に心室に強い刺激が加えられるとVTやVFなどの致死性不整脈が誘発されることがあります。
これをR on Tと呼びますね(ペースメーカではSpike on T)。
さいごに
以上で、活動電位に関わる内容でした。
筆者自身も執筆しながら勉強できましたし、内容の濃いものにできたのではないかと思っています。
さて、私も思ったことがあるのですが、「Cl-やMg2+は活動電位ではどのような役割があるのか」といったところです。番外編として、Cl-やMg2+について執筆を考えております。ご期待ください。












コメント